 |
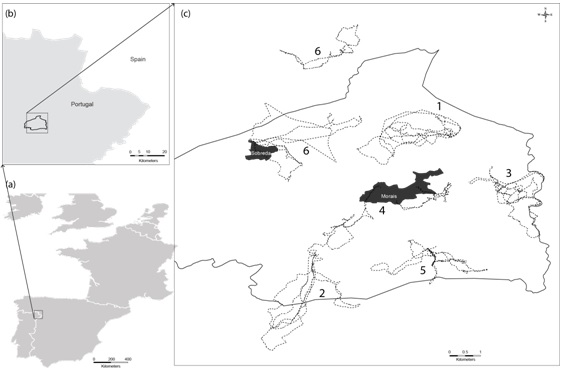
| Figure 1. Map of the location of grazing circuits of the six herbs where 1-3: goat herds and 4-6: sheep herds |
| Livestock Research for Rural Development 28 (5) 2016 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
The objective of this study was to compare the feeding strategies between sheep and goat rearing by shepherds in the Mediterranean mountain area of northeast Portugal. For this purpose, grazing itineraries, diet selection, diet overlap and the diet diversity of different flocks of sheep and goats were studied. The flocks were monitored along specific grazing circuits every three months. Grazing itineraries were recorded by means of a hand global positioning system (GPS). Diet composition was determined by direct observation method in pre-set intervals of 15 minutes during the day. For each observation point, herbaceous communities, shrubs and tree species consumed were recorded. Diet selection was estimated by Krueger’s preference index and the degree of overlap between diets was estimated using the Kulczynskis similarity index.
The diet of goats had a significantly higher content of shrubs (27%) and trees species (24%) than sheep (6% and 10% respectively). Sheep diet comprised 84 % of herbaceous plants, whereas this group only represented 49 % in goat diet. Erica sp. and Rubus sp. were the shrubs with the highest preference index for goats. Both animals avoided Cytisus multiflorus in winter and spring, and Cistus ladanifer in summer. Goats showed a higher preference for Fraxinus excelsior and Quercus rotundifolia. Goat diversity of forage on offer and animal diet was significantly higher than on the sheep diet. Plant species diversity in sheep and goat diets significantly increased in autumn compared to spring. The average of diets’ overlap was higher during the winter and lower in summer. The length of grazing itineraries and diet diversity reflects the level of dependence on natural resources and consequently it´s level of extensification.
Keywords: diet diversity, diet overlap, diet selection, small ruminants
Rangelands are a terrestrial natural resource with great ecological, economic and social importance for many regions of the world (Jouven et al 2010; Karimzadegan et al 2007). Although they are seen as unproductive lands, rangelands constitute the basis of various activities that are often compatible with each other. For instance, they offer the opportunity to carry out outdoor recreational activities (Castro 2012), and give shelter to biodiversity conservation (Bugalho and Abreu 2009). Rangelands occupy 32% of the total land surface of Portugal (ICNF 2013), and together with the agricultural by-products, they are the basis of livestock feeding systems of mountain areas in this region.
On the other hand, forestland use occupies about 68% of the territory (semi-natural grasslands and shrublands: 43%; woodlands: 25%) (ICN 2006). Q. rotundifolia (11% of land use forest) and Q. suber (10% of land use forest) are the most important species, followed by others, such as Q. faginea and Juniperus oxycedrus.
Natural pastures, forages, fodder legumes, woody species, and crop residues are consumed by sheep and goat flocks in a pastoral system based on grazing itineraries. Throughout the year, the consumption of these feed resources differs according to several factors such as forage availability and accessibility, plant phenological stage (Moore and Jung 2001), plant content in anti-nutritional or toxic compounds (Bryant et al 1992), the physical defences of plant itself, animals’ species and breed, and the nutritional and physiological state of the animals.
The Mediterranean climate is characterised by hot dry summers and cool wet winters, and during favourable seasons (autumn and spring), pastures are available in higher quantities and show good nutritional quality, whereas the dry season’s pastures have poor nutritional quality with high fibre and low protein contents, which often results in seasonal weight loss of livestock (Lamy et al 2012). In the summer and winter, it is usually necessary to provide additional resources such as forage, hay or grains to feed the flocks. Fodder trees and shrubs are common components of Mediterranean vegetation and often fill the feed gap of the summer as well as the winter when evergreen (Kebaili and Papanastasis 2005). Furthermore, the seasonal availability of their sprouts, leaves, flowers and fruit may moderate the variations in the total amount of edible matter (Ammar et al 2004).
In the north of Portugal, rangelands are grazed by flocks of sheep and goats, always guided by a shepherd (Castro and Gomez Sal 2016); usually there is no overlap in the use of grazing territory (Castro et al 2004). Sheep graze first on meadows and forage areas around the homestead, while goats, however, use woodlands in the remote areas of the hamlet. Hence, sheep and goats are commonly classified as different feeding types, the former as mixed feeders and the latter as browser (Hoffman, 1989; NRC 2007). Despite this, faced with a scarcity of feed resources particularly in the dry season, all animal species fall back on browse species (Sanon et al 2007).
Currently, improving the management of the available rangelands in a region is a high priority, owing to their potential for commercial livestock production, their important social function regarding fixing the population and maintaining traditions, and the interest in decreasing wildfire risk. Despite the fact that sheep and goats exhibit different feeding habits, when adequately managed they can produce excellent results for wildfire prevention as they can control woody biomass growth (Papanastasis 2009; Ruíz-Mirazo and Robles 2012; Lovreglio et al 2014).
Assessing the behaviour patterns of small ruminants in extensive pastoral systems will provide a baseline of information to make management decisions and increase the efficiency of grazing systems in theses areas. The objective of this study is to compare the grazing itineraries, the diet selection, the overlapping levels and the diet diversity of different flocks of sheep and goats on Mediterranean mountain rangelands.
The experiment was carried out in the Morais region (N41° 29' 23" W6° 46' 44"; 600 m above see level), northeast of Portugal (Nature network 2000, PTCON0023, Fig. 1). The climate is sub-humid Mediterranean. The mean annual temperature is 14.6 ºC and the mean rainfall is 636 mm, which occurs mainly from October until May (INMG 1991). Geomorphologically, it is characterised by an undulated plateau (mean altitude of 600 m), cut by two rivers—the Azibo and Sabor (Pacheco and Van der Weijden 2002). It is one of the most representative areas of ultramafic rocks, and one of the largest continuous units of serpentinite in Portugal. Soils are shallow and poor in nutrients, and frequently rich in heavy metals such as nickel and copper. Perennial sclerophyllous woodlands with changeable combinations of Fagaceae (Q. suber L., Q. rotundifolia Lam., Q. faginea Lam. and Crupressaceae species Juniperus oxycedrus L.) constitute the natural potential vegetation of this region (Sequeira et al 2010).
|
| Figure 1. Map of the location of grazing circuits of the six herbs where 1-3: goat herds and 4-6: sheep herds |
Subsistence farming is the dominant type of agriculture. Winter cereals (wheat and rye) and spring crops (maize, potatoes or vegetables) are produced in small areas. Also, perennial crops, mainly olive groves and some vineyards are very common in the landscape. A very significant part of the arable land of the village is cultivated with fodder plants, mainly barley, oats, fodder sorghum and other forages.
Livestock animals are raised by landless farmers or smallholders. Rangelands are extensively used by livestock and a sheep and goat rearing system is supported by a grazing circuit network over all the non-fenced or uncultivated village territory in which natural resources are constrained and people and their animals adopt strategies to meet these constraints. As a result, grazing circuits and consumed resources strongly differ throughout the year and throughout the territory, in other words between seasons and animal species.
Six flocks, three of sheep (Churra da Terra Quente breed) and three of goats (Serrana breed), were randomly selected from the flocks identified in the study area. The flocks, led by their shepherd, use several grazing circuits, depending on season and availability of resources. Along these grazing circuits, animals cross a land use mosaic composed by different feed resources: meadows, annual and perennial crops, rangelands and forestlands. The flocks were monitored every three months for a year in order to determine their grazing circuits and diet composition. Field observations were made in September (autumn) 2010, January (winter), April (spring), and July (summer) 2011. The flocks set out daily for pasture on schedules that differ with the season, abundance of resources and animal species. The flocks return to their stables every day. In the summer, flocks of sheep set out before sunrise and go back after sunset. Back home, animals receive straw and hay during the scarcity periods (winter and summer). The flocks have a variable size (110, 120, 130: goats/flocks; and 170, 170, 340: sheep/flocks). The live weight of goats and sheep ranged from 25 to 40 kg and 55 to 60 kg, respectively. Livestock are raised for meat production and the management is considered to be extensive.
To determine the length and duration of grazing circuits, each herd was recorded one day per season by means of a hand global positioning system (GPS). GPS data comprised time, geographical position and land cover of 24 herd itineraries in total.
Plant species as well as the herbaceous communities were previously identified along grazing areas of the Morais territory so that they would be recognised correctly. At each observation point, herbaceous communities, shrubs and trees species consumed were recorded.
Five herbaceous communities were considered: i) herb (Cv), mainly composed of spontaneous grasses and forbs; ii) cereal-fallow (Ba); iii) meadow (M); iv) cereal stubble (Re); and v) other forage (Ot). The availability of each species or vegetal group was evaluated by visual estimation of plant species cover as described by Torrano and Valderrábano (2005) by two observers (at each observation point, where feeding activities were recorded).
Diet selection was determined by the direct observation method, following the methodology of Meuret et al (1985) with slight modifications. The scan-sampling method proposed by Altmann (1974), consisting of systematic sampling at pre-set time intervals and regular records, was followed in the present work. At each observation point, two observers recorded the activity of 10 animals randomly chosen in a quadrant (i.e. N, NE, E, SE, S, SW, W, NW). The set of animals and quadrant changed every 15 minutes. If animals were feeding, the plant species (or “facies”) consumed were recorded, as proposed by Agreil and Meuret (2004). Data collection was performed from points close to the groups of animals found. Once animals were familiar with human presence, the observer followed the flocks very closely and the grazed species could be identified without any difficulties. The total number of recordings per day varied between 23 and 48 according to animal species and seasons. During the summer, when temperatures were very high, flocks of sheep were also monitored at night because they graze after sunset.
Plant species as well as the herbaceous communities were previously identified along grazing areas of the Morais territory so that they would be recognised correctly. At each observation point, where feeding activities were recorded, herbaceous communities, shrubs and trees species consumed were recorded.
The availability of each species or vegetal group was evaluated by visual estimation of plant species cover as described by Torrano and Valderrábano (2005) by two observers (at each observation point, where feeding activities were recorded). It was assumed that the number of recordings of a plant species reflects the proportion of that plant in the diet. The percentage of each plant in the diet was calculated by the ratio between the numbers of records in relation to total number of recordings foraging activity per observation day (Castro 2014).
Diet selection was estimated by Krueger’s preference index (Krueger 1972) using the following equation:
RPi = Σk=1,n (Dik/RAik)/n
where RPi is the mean preference ratio over n areas; Di is the percentage of species i in the diet, and RA i is the percentage of species i in the area. Values of RP greater than 1 indicate preference, while values less than 1 indicate avoidance. The preference index was calculated for each sampling season (spring, summer, autumn and winter).
Diversity of forage on offer and animal diet were estimated according to the Shannon-Weaver index (Pielou 1975) using the following equation:
H´ = - Σ pi log10Pi
where Pi is the proportion of the total diet or forage in offer belonging to the ith species.
The degree of overlap between the diets was estimated using the Kulczynskis similarity index (Oosting 1956) using the following equation:
KSI = [Σ2W/Σ (a + b)] ×100
where W is the lesser percentage of a common component in the two diets to be compared and (a + b) is the sum of percentages of components of the two diets being compared. This index ranges from 0 (no overlap) to 1 (total match). KSI was estimated in each sampling season.
Grazing itineraries (length and duration), diet selection, plant diversity and diet overlaps were analysed via ANOVA, with the objective to compare the means between this variables and determines whether any of those means are significantly different from each other, following the model: Yijklm = μ + SPi + SSj + ESk + εijk, where Yij is the studied variable; μ the variable mean; SPi: plant categories (herbaceous, shrubs and trees); SSj: sampling season; ESk experimental site; and ε ijk the error. The LSD (least significant difference) test was used to make subsequent pairwise comparisons after a normalisation test (α = 0.05), but only when factors were significant according to the results of the ANOVA analyses. The statistical software package SAS (2001) was used for all analyses.
On the other hand, preference or rejection of vegetation component was estimated expressed as a function of its occurrence in the diet and its availability to the animals. Availability in the present study was estimated as the vegetation composition accessible to the animals weighted for the area of the different range or vegetation types and the time spent in each type.
Sheep spent more daytime on grazing itineraries than goats (P < 0.05) independently of the season, in average 504 min d-1 compared to 409 min d-1 (Table 1). The greatest differences were found in summer and spring where duration grazing itineraries of sheep were higher than goats, 188 min and 102 min, respectively. On the other hand, the length of the grazing itinerary was significantly longer in autumn compared to the other seasons (P < 0.001).
|
Table 1. Duration of the grazing per day and distances walked per day by season in goats and sheep (mean ± SEM). |
||||||||
|
Duration of |
Length of grazing |
p value |
||||||
|
Length |
Duration |
|||||||
|
Goats |
Sheep |
Goats |
Sheep |
Animal |
Animal*Season |
Animal |
Animal*Season |
|
|
Autumn |
528a
|
562a
|
7.02a
|
10.1a
|
0.66 |
0.02 |
0.18 |
0.04 |
|
Winter |
346b
|
402b
|
6.14a
|
5.07b
|
||||
|
Spring |
388b
|
490ab
|
5.16a
|
4.00c
|
||||
|
Summer |
376b
|
564a
|
6.13a
|
6.27b
|
||||
| abc Different letters indicate significant differences between seasons for the same animal species | ||||||||
The percentage of shrubs and tree species in the diet of goats was significantly higher (P < 0.01) than in that of sheep, while the proportion of herbaceous was lower in the diet of goats (P < 0.001). The presence of tree species in sheep and goat diets increased significantly in autumn (P < 0.05) (Fig. 2). Shrubs ingestion tended to increase during summer and winter in goats, and in the case of sheep, it is highest in winter.
 |
|
Figure 2.
Seasonal percentage of herbaceous, shrubs and trees in the diet of
goats and sheep. Different letters indicate significant differences in the same vegetation type between seasons |
The number of plant species or plant groups in the diet ranged between 10 to 20 and 7 to 13 in goats and sheep, respectively, with the maximum diversity observed in autumn and the minimum in spring (Table 2). Forbs and grasses (31%), Sorghum sp. (16%) and Q. rotundifolia (13%) were the most consumed groups in the case of goats, while meadow (34%), herb (25%) and stubble (10%) were the most consumed in the case of sheep.
|
Table 2. Percentages of the total species/season obtained by the direct observation of goats and sheep grazing. Cod: Species code; Aut: Autumn; Win: Winter; Spr Spring; Sum: Summer |
||||||||||
Species |
Goats |
Sheep |
||||||||
|
Cod |
Aut |
Win |
Spr |
Sum |
Aut |
Win |
Spr |
Sum |
||
|
Herbaceous |
Avena sp. |
Av |
- |
- |
- |
- |
6.80 |
- |
- |
10.6 |
|
Hordeum vulgare |
Hv |
- |
- |
- |
- |
- |
1.80 |
- |
- |
|
|
Sorghum sp. |
So |
24.5 |
- |
- |
7.80 |
8.00 |
- |
- |
- |
|
|
Vigna unguiculata |
Vu |
- |
- |
- |
- |
3.70 |
- |
- |
- |
|
|
Forbs + Grasses |
Cv |
14.4 |
41.9 |
57.2 |
11.2 |
11.9 |
46.2 |
39.2 |
0.60 |
|
|
Other forage |
Ot |
0.80 |
- |
0.50 |
3.20 |
- |
- |
- |
7.90 |
|
|
Cereal fallow |
Ba |
3.20 |
- |
1.70 |
- |
- |
- |
12.5 |
5.00 |
|
|
Meadow |
M |
2.60 |
- |
9.30 |
15.3 |
34.8 |
19.7 |
42.2 |
39.5 |
|
|
Stubble |
Re |
1.20 |
- |
- |
- |
12.3 |
10.6 |
- |
19.0 |
|
|
Shrubs |
Cistus ladanifer |
Ci |
8.00 |
18.8 |
5.70 |
7.20 |
0.20 |
0.10 |
0.50 |
1.90 |
|
Cistus psilosepalus |
Cp |
1.40 |
- |
- |
- |
- |
- |
- |
- |
|
|
Cytisus multiflorus |
Cm |
0.40 |
6.90 |
11.1 |
- |
- |
10.3 |
5.10 |
- |
|
|
Cytisus scoparius |
Cs |
- |
- |
- |
1.90 |
- |
- |
- |
- |
|
|
Daphne gnidium |
Dg |
- |
0.40 |
- |
- |
- |
- |
- |
- |
|
|
Erica sp. |
Er |
2.00 |
5.30 |
- |
5.90 |
- |
- |
- |
3.20 |
|
|
Genista hystrix |
Gh |
- |
1.90 |
1.60 |
6.70 |
- |
- |
- |
- |
|
|
Lavandula pedunculata |
Lv |
0.40 |
2.50 |
0.90 |
- |
- |
- |
- |
0.90 |
|
|
Rubus sp. |
Ru |
4.20 |
0.90 |
1.80 |
10.7 |
- |
0.20 |
0.30 |
0.20 |
|
|
Alnus glutinosa |
Al |
4.70 |
- |
- |
5.30 |
- |
- |
- |
- |
|
|
Trees |
Crataegus monogyna |
Cr |
0.60 |
- |
- |
- |
- |
- |
- |
- |
|
Fraxinus excelsior |
Fr |
3.80 |
- |
- |
3.00 |
3.10 |
- |
- |
- |
|
|
Juniperus oxycedrus |
Jc |
0.60 |
- |
- |
- |
- |
- |
- |
- |
|
|
Olea europaea |
Ol |
1.40 |
18.3 |
- |
2.40 |
10.2 |
6.90 |
- |
9.10 |
|
|
Quercus faginea |
Qf |
- |
- |
- |
- |
0.5 |
- |
- |
- |
|
|
Quercus rotundifolia |
Qr |
21.0 |
2.90 |
10.0 |
17.8 |
4.30 |
4.10 |
0.20 |
1.80 |
|
|
Quercus suber |
Qs |
2.50 |
- |
- |
- |
1.10 |
- |
- |
- |
|
|
Ulmus glabra |
Ul |
0.70 |
- |
- |
1.40 |
0.20 |
- |
- |
- |
|
|
Total |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
100 |
||
In the case of goats, species such as Cistus psilosepalus and Q. suber were consumed only in the autumn, whileCytisus scoparius and Daphne gnidium were consumed only in the summer and winter, respectively. In the case of sheep, Sorghum sp., Vigna unguiculata, Fraxinus excelsior, Q. faginea, Q. suber and Ulmus glabra were consumed only in autumn, while Erica spp. and Lavandula pedunculata were present in the diet of sheep only during the summer. Furthermore, some species such as Cistus ladanifer, Q. rotundifolia and Rubus sp. were consumed by both animals during all seasons with the exception of Rubus sp. in the case of sheep in autumn.
The comparison results between species selection versus species rejection by goats and sheep during the grazing period can be seen in Fig. 3. Herb and Sorghum sp. showed to be strongly consumed by goats in spring (Fig. 3E) and autumn (Fig. 3A), respectively. Regarding herb, the results showed that goats decreased its consumption in the other grazing seasons. In the case of sheep, herb was selected mainly in autumn as well as stubble (Fig. 3B), and it was less consumed in spring (Fig. 3F) and summer (Fig. 3H). Meadows were selected across the seasons by sheep. Shrub species such as Cistus ladanifer were strongly selected in winter by goats (Fig. 3C), but its selection decreased in the other seasons. Sheep weakly selected the previous species in all seasons. The same result was obtained in the case of Cytisus multiflorus for both animal species. Regarding tree species, the results showed that Q. rotundifolia was selected by goats in all seasons of the year, mainly in autumn (Fig. 3A), and summer (Fig. 3G), while by sheep, only in autumn (Fig. 3B) and winter (Fig. 3D),. On the other hand, Olea europaea was shown to be consumed proportionally above their abundance in the case of sheep and of low consumption by goats.
 |
| Figure 3. Species selection vs. rejection by goats and sheep in autumn, winter, spring and summer.
Note: *: to facilitate the graphical representation of the data they were divided into three. Code species can be seen in Table 2 |
Finally, herbaceous, shrub and tree species in the diet of goats and sheep were compared to their respective abundance in the study area in order to determine the individual species preference (Fig. 4).
 |
| Figure 4. Seasonal goats and sheep Krueger´s preference index for herbaceous, shrubs and trees species |
In the case of herbaceous, sheep showed a higher preference (RPI > 2) for stubble in winter, while both animal species showed a weak preference (1< RPI < 2) for it and Sorghum sp. in autumn. The shrub species with the highest preference index (both species in summer) were Erica spp. for sheep, and Rubus sp. for goats. Cytisus multiflorus was avoided during winter and autumn by both animal species. On the other hand, goats showed a higher preference for Fraxinus excelsior in summer, while both animal species avoided it in autumn. Other tree species such as Q. rotundifolia were highly preferred by goats in autumn and spring, whereas sheep showed little preference for them in autumn, and avoided them during the spring.
Diversity of forage on offer and animal diet of goats’ diversity was significantly higher than that of sheep (P < 0.01) (Fig. 5). On the other hand, plant species diversity in the diet of both animal species significantly increased in autumn in relation to spring ( P < 0.05).
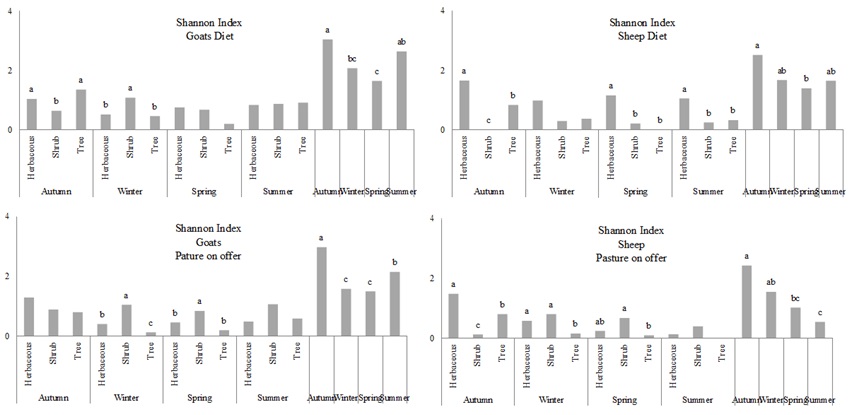 |
| Figure 5. Shannon index for diet and forage on offer in goats and sheep. Different letters indicate significant differences between vegetation in the same season |
Plant species availability for goats was lower during the winter and spring (P < 0.001) while in the case of sheep, higher values were found in autumn compared to spring and summer, and in winter compared to summer (P < 0.01).
Diet/Diet overlaps
The observed overlaps reflect seasonal influences as animals shift diet focus (Fig. 6). In the case of herbaceous species, the overlap was significantly higher in spring (P < 0.01; 0.35), and lower in summer (0.18). In the case of shrub species, the highest values were found in spring (0.22), and the lowest in autumn (0.01). Regarding tree species, the degree of overlap was lower during spring (0.04) and summer (0.12) and higher during winter (0.68). The average overlap of diets was greater during winter (0.36) and lower in summer (0.16).
 |
|
Figure 6.
The degree of overlap between diets estimated
using Kulczynski’s Similarity Index (KSI). Different letters indicate significant differences between seasons for the same vegetation type |
The duration of the daily grazing itineraries is affected by the availability of food resources and the kind of livestock production system, as well as by environmental limitations that change behaviour patterns of ruminant animals. The length of the daily grazing itineraries of goats was similar to values reported by Pacheco (2000) (10 km/d) in Serra da Penada (NE of Portugal); and lower than the values reported by Schlecht et al (2009) in northern Oman, due to the different ecological conditions (Total annual precipitation of 100-340 mm; Annual average temperature 18.1 ºC with a minimum of 3.6 ºC and a maximum of 36.3ºC). In the case of sheep, our results showed higher length values than those found by Castro (2004) in the northeast of Portugal due to the absence of nocturnal grazing. The results of the present study showed significant seasonal differences in the itineraries length in both animal species due to the changes in forage availability. Thus, the longer duration of itineraries was found in autumn due to the scarcity of forage in contrast to the situation in spring, when the availability of forage increases, and therefore, the length of itineraries was reduced. On the other hand, goat and sheep grazing times observed in this study were similar to those reported in previous studies (480-630 min/day: Lynch et al 1992; Bojkovski et al 2006; Ferreira et al 2013). In our case, excluding the flocks of sheep in summer, grazing time was limited to daylight periods, coinciding with the activity patterns for sheep and goats under extensive conditions as generally described in literature (Askins and Turner 1972; Lu 1988). Furthermore, the the results showed that goats spent less time grazing than sheep due to their lower overall dry matter intake associated with their lower body weight as reported by Bonanno et al (2008). On the other hand, our results showed that the grazing time varied between seasons, being the highest in autumn coinciding with the lowest forage availability and when the days are shorter, and the lowest in winter coinciding with the lowest average temperature in the area of the study. In the case of sheep, it should be pointed out that the higher time of grazing was also observed in summer where the average temperature in the area increased. Hence, on hot days, sheep grazing occurred early in the morning and at night, while the activity ceases and remains inactive during the hot hours of the day, increasing the time of grazing. Compared to goats, it is known that sheep are particularly sensitive to high temperatures and solar radiation, and hence, nocturnal grazing is more beneficial for sheep, contributing to improving body condition, reducing heat stress and increasing forage intake and production (Marai et al 2007).
According to our results, sheep behaved as typical grazers selecting more herbaceous species in their diet. Although the contribution of herbaceous species in the diet of goats was also important, shrubs and tree species represented the main component in the diet of goats as previously observed in other studies (Allan and Holst 1996; Grant et al 1987; Celaya et al 2007; Osoro et al 2013). The results showed that herbaceous resources dominated the diet of both animals during spring (94% and 69% in sheep and goats, respectively) when these resources showed higher protein content and highest digestibility (Fernández-Núñez et al 2012). During spring, the most important herbaceous components in the diet of both animals were herb and meadow. Despite the high percentage of herb in the diet of sheep, it is important to highlight that this resource was consumed below their proportional abundance in all seasons with the exception of autumn when the availability of this resource decreases. Regarding meadow, it is known that the growing season in the Mediterranean area limits the production of meadows to spring and autumn (Pires and Moreira 2000). Moreover, the quality of meadows is low during summer. In that period, the grass in meadows is dry and depleted, and it is customary that shepherds lead their flocks for grazing to other resources such as stubble. However, our results showed that meadow continued to be an important component in the diet of both animal species during the summer, while stubble was the second component in the diet of sheep during this season. This result could be explained by the fact that flocks were monitored at the beginning of summer, after the hay was cut, when the nutritive value of meadows remains relatively high.
It is important to highlight that sheep exhibited a strong preference for stubble during the winter (Table 2) despite due to the large amount of indigestible stem material that reduces their nutritive value during this season. This result is in contrast with other authors who reported higher preference for this type of resource during the summer and autumn (Robertson 2006). One hypothesis that could explain such findings is that while the ground is still wet from the night’s dew, the shepherds lead their flock to pastures sparsely covered with plants, and in our case, stubble where sheep are attracted to graze the cereal grain that remain on the soil surface. When the rising sun has dried the dew, the flock moves on to more nutritive pastures. >
It has been established that the diet composition of goats and sheep changes seasonally, according to plant species availability, their phenological development and their nutritive value (Fajemisin et al 1996; Provenza et al 2003; Utsumi et al 2009; Basha et al 2012). The results of the present study showed that shrubs species were always present in the diet of goats. For example, species such as Cistus ladanifer and Rubus sp. were present in the diet of goats in all seasons. Previous research showed that Rubus sp. provided excellent quality forage suitable for livestock (McGregor 1992). This author points out that the consumption of this genus in the summer and autumn when pastures have low levels of digestibility would be superior to providing by hay. Goats and other animals such as roe deer (González-Hernández et al 2003) and horse (Rigueiro et al 1998) are effective at removing the above ground parts of this genus, while pigs have been shown to be very effective at digging up and consuming the roots (Rodríguez-Estévez et al 2009). On the other hand, the diet of goats was dominated by shrub species in winter and summer but their consumption decreased in autumn and spring. It is known that goats are deemed as opportunistic feeders, eating the best that is available to them, while sheep are deemed as grazers (Kawas et al 2010; Egea et al 2014). Thus, goats increased the consumption of herbaceous species in seasons in which the nutritional values of herbaceous species in the Mediterranean area are high (autumn and spring) (Pires and Moreira 2000). Moreover, most of the shrub species present in the area of the study showed a low quality (low protein and high fibre) during the autumn (Castro and Fernández-Núñez 2014) and goats avoided them (e.g. Cytisus scoparius, Genista hystrix, Cytisus multiflorus). One exception to these results can be seen in the case of Cytisus multiflorus. The percentage of this shrub species in the diet of goats increased during the spring, coinciding with the absence of tannins (González-Hernández et al 2003).
Unlike for shrubs, the presence of tree species significantly increased in both animal species in autumn, contributing to an increase in the diet diversity values. These results can be explained; when resources are limited, a change in feeding strategy from specialist to generalist occurs due to the necessity of maintaining nutrient intake. Goats showed higher intakes than sheep for all tree species with the exception of Q. faginea andOlea europaea. These results can be explained by the fact that in the presence of other tree species, mainly Q. rotundifolia and Fraxinus excelsior, goats had chosen these tree species at the expense of Olea europaea (Table 1). However, it is important to highlight that both animals showed a higher consumption of Olea europaea during the winter. Olive leaves provide half of the energy and amino acid requirements of sheep and goats at the maintenance level (Molina-Alcaide and Yáñez-Ruiz 2008) and they have traditionally been used in the Mediterranean area as alternative feeds for sheep and goats during periods of scarcity (winter). Another important component of the diet of both animal species, but especially of goats, was Q. rotundifolia during autumn and winter. In these seasons, sheep and goat flocks eat acorns. It is known that oak acorns have potential nutritive values for ruminant animals since acorns have high starch, organic matter and metabolisable contents but a low level of condensed tannin contents (Al Jassim et al 1998; Rodríguez-Estévez et al 2008; Kaya and Kamalak 2012). Sheep and goats exhibit different levels of tolerance to the effects of tannins extracted from Quercus spp. leaves (Narjisse et al 1995). Furthermore, under field conditions goats are able to consume larger amounts of tannin-rich Quercus leaves than sheep (Min and Hart 2003). On the other hand, condensed tannins concentration decreased with maturation in Quercus spp. leaves (Kamalak et al 2004). All of these reasons could help to explain the higher selectivity showed by goats compared to sheep for Q.rotundifolia during the spring and summer in the study area.
Moreover, goats showed a higher preference for Fraxinus excelsior in the summer as previously reported by Papachristou and Papanastasis (1994) because this species’ foliage supplied sufficient digestible protein and energy to meet the production requirements of these animals (Papachristou and Nastis 1996; Pereira et al 2008).
The selective behaviour of animals changes from season to season according to the available resources. This different behaviour is shown by the partial replacement of vegetation resources in seasons in which they were scarce and other resources were available. For these reasons, the diet overlap between goats and sheep for each kind of resource changes from season to season (Fig. 6). Thus, the fact that herbaceous species were highly abundant, and therefore, easily accessible to livestock in spring in relation to the other seasons, would explain the highest diet overlaps found in this type of vegetation in spring (Fig. 6). Regarding trees species, the highest overlap was found in winter when goats and sheep showed higher preference for Q. rotundifolia and Olea europaea for the reasons explained above. However, goats and sheep showed low diet overlap in the case of shrub species, suggesting little competition for this type of vegetation. The greatest diet overlap for shrubs occurred in summer when the availability and quality of herbaceous species were lower and their desirability decreased.
The socio-economic importance of rangelands is widely recognised but it could be constrained by inefficient use of pastoral systems and poor feeding management. The results obtained in this work showed a seasonal effect on the utilisation ability of resources by goats and sheep that should be taken into account when designing management practice in the area. This variation in preference for some plant species could be related to modifications in their nutritive value or secondary metabolism content (e.g. tannins). Many species with appreciable amounts of tannins, such as the genus Cistus> sp., Erica sp., Quercus sp. sp., were present, above all, in the diet of goats. However, goats reduce their consumption of most of these species in the spring, when the availability and nutritive value of herbaceous species increase in the study area. This result contributes, on one hand, to an increase in the diet overlap for herbaceous resources in spring, and on the other hand, to an increase in the shrubs accumulation, and therefore, increases the risk of wildfires in the area. However, shepherds need to provide supplementary forage to feed the flocks in summer and winter when the availability of herbaceous species decreases in the study area. Taking into account that when shepherds control their flocks on a grazing circuit, their strategy for utilisation of resources influences the animal’s behaviour. This makes it possible to make some management recommendations for spring grazing in the case of goats: i) goats could be guided by the shepherd to shrub areas to promote their consumption and reduce the intake of herbaceous resources that could be used for hay during the scarce periods; ii) increase the size/number of the flocks, which could increase the shepherd’s revenues, taking into account that the ability of the shepherd to control the animals is a determining factor for the maximum size of the flocks. The maximum size of the flocks is approximately 150 goats in the study area; iii) mixed grazing by goats and sheep using goats as support thus influencing the consumption of more woody species by sheep as this type of animal has a more passive foraging behavior.
This study was funded by the Novo Norte Programme / Câmara Municipal de Macedo de Cavaleiros. The authors would like to thank the shepherds of goat and sheep flocks in Morais village for their collaboration and field assistance and in particular to José and Américo Peso, José Martins, Teodoro Major, Rodrigo e Baldomero. The authors would also like to thank Daniel Saraiva for his field assistance and José Castro for his drawing assistance.
There are no conflicts of interest.
Agreil C/span> and Meuret M 2004 An improved method for quantifying intake rate and ingestive behaviour of ruminants in diverse and variable habitats using direct observation. Small Ruminant Research 54: 99-113.
Al Jassim R A M, Ereifej K I, Shibli, R A and Abudados A 1997 Utilization of concentrate diets containing acorns (Quercus aegilps and Quercus coccifera) and urea by growing Awassi lambs. Small Ruminant Research 29: 289-293.
Allan, C J and Holst P J 1996 The ecological role of the goat in maintaining pasture and range. In: VI International Conference on Goats. International Academic Publishers: Beijing, China, pp. 427-435
Altmann J 1974 Observational study of behaviour: sampling methods. Behavior 49: 227-267.
Ammar H, López S, González, J S and Ranilla M J 2004 Seasonal variations in the chemical composition and in vitro digestibility of some Spanish leguminous shrub species. Animal Feed Science and Technology 115: 327-340.
Askins G D and Turner E E 1972 A behavioral study of angora goats on West Texas range. Journal of Range Management 25: 82-87.
Basha N A D, Scogings PF, Dziba L E and Nsahlai I V 2012 Diet selection of Nguni goats in relation to season, chemistry and physical properties of browse in sub-humid subtropical savanna. Small Ruminant Research 102: 163-171.
Bojkovski D, Stuhec I and Kompan D 2006 Behaviour of sheep in three different types of paddock in karst region of Slovenia. Acta Agraria Kaposváriensis 10: 159-167.
Bonanno A, Fedele V and Di Grigoli A 2008 Grazing management of dairy goats on Mediterranean herbaceous pastures. In: A. Cannas and G. Pulina (Eds) Dairy Goats Feeding and Nutrition. CABI: London, UK, pp. 189-220.
Bryant J P, Reichardt P B and Clausen T P 1992 Chemically mediated interactions between woody plants and browsing mammals. Journal of Range Management 45 (1): 18-24.
Bugalho M N and Abreu J M 2009 The multifunctional role of grasslands. C. Porqueddu and Tavares de Sousa M.M. (Eds.) Sustainable Mediterranean grasslands and their multi-functions Zaragoza. CIHEAM / FAO / ENMP / SPPF. Option méditerranéennes Serie A Séminaires Méditerranéens; n. 79. pp. 25-30. http://om.ciheam.org/om/pdf/a79/00800611.pdf
Castro J 2012 A fruição do silvestre: o caso do monte Morais. Porto: Faculdade de Ciências. Dissertação de Mestrado em Arquitetura Paisagista. Universidade do Porto, Porto, Portugal.
Castro M 2004 Análisis de la interacción vegetación-herbivoro en sistemas silvopastorales basados en Quercus pyrenaica. Tesis Doctoral, Universidad de Alcalá, Alcalá de Henares, Spain.
Castro M, Castro J F and Gómez-Sal A 2004 Quercus pyrenaica Willd. woodlots and small ruminant production in Northeast Portugal. In: S Schnabel and A Ferreira (Eds) Sustainability of agrosilvopastoral systems, dehesas, montados. Catena: Reiskirchen, Germany, pp. 221-229.
Castro M and Fernández-Núñez E 2014 Small ruminants as a fire management tool in a Mediterranean mountain region. In: JHN Palma and A Chalmin (Eds) Second European Agroforestry Conference: integrating science & policy to promote agroforestry practice. EURAF: Germany, pp. 48-51.
Castro M and Gomez Sal A 2016 Bosquetes de Quercus pyrenaica Willd. en sistemas de pastoreo dirigido. Situación actual y perspectivas en Trás-os-Montes (Portugal). M.R. Mosquera-Losada, G. Moreno and A. Rigueiro-Rodríguez (Eds) Sistemas Agroforestales en la Península Ibérica, Spain (In press).
Celaya R, Oliván M, Ferreira, L M M, Martínez A, García, U and Osoro, K 2007 Comparison of grazing behaviour, dietary overlap and performance in non-lactating domestic ruminants grazing on marginal heathland areas. Livestock Science 106: 271-281.
Egea V, Allegretti L, Paez Lama S, Grilli D, Sartor C, Fucili M, Guevara J C and Passera C 2014 Selective behavior of Creole goats in response to the functional heterogeneity of native forage species in the central Monte desert, Argentina. Small Ruminant Research 120: 90-99.
Fajemisin B, Ganskopp D, Crus R and Vavra M 1996. Potential for woody plant control by Spanish goats in the sagebrush steppe. Small ruminant research, 20: 99-107.
Fernández-Núñez E, Pires J M, Fernandes A, Pires J, Aguiar, C, Galvão L and Moreira N 2012. Grazing regimes and fertilisation rates: effects on dry-matter yields, crude protein content and digestibility of meadows in the Northeast of Portugal. In: P Goliński, M Warda and P Stypiński (Eds) Grassland science in Europe, 17: 311-313.
Ferreira L M M, Celaya R, Benavides R, Jáuregui B M, García U, Santos A S, García, R R, Rodrigues M A M and Osoro K 2013 Foraging behaviour of domestic herbivore species grazing on heathlands associated with improved pasture areas. Livestock Science 155: 373-383.
González-Hernández M P, Karchesy J and Starkey EE 2003 Research observation: hydrolysable and condensed tannins in plants of northwest of Spain forest. Journal of Range Management 56: 461-465.
Grant S A, Torvell L, Smith H K, Suckling D E, Forbes T D A and Hodgson J 1987 Comparative studies of diet selection by sheep and cattle: blanket bog and heather moor. Journal of Ecology 75: 947–960
Hoffman R R 1989 Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78: 443–457.
ICN 2006 Plano Sectorial Rede Natura 2000. Sitio Morais (PT CON0023). Instituto de Conservação da Natureza. Lisboa. Portugal.
ICNF 2013 Áreas dos usos do solo e das espécies florestais de Portugal continental. Resultados preliminares. Instituto da Conservação da Natureza e as Florestas. Lisboa. Portugal.
INMG 1991 O clima de Portugal. Normas climatológicas da região de Trás-os-Montese Alto Douro e Beira Interior correspondentes a 1951-1980. Instituto Nacional Metereologia Geofisica. Lisboa. Portugal.
Jouven M, Lapeyronie P, Moulin C H and Bocquier F 2010 Rangeland utilization in Mediterranean farming systems. Animal 4 (10): 1746–1757.
Kamalak A, Canbolat O, Ozay O and Aktas S 2004 Nutritive value of oak (Quercus spp.) leaves. Small Ruminant Research 53: 161-16.
Karimzadegan H, Rahmatia M, Dehghani Salmasi M, Jalali R and Shahkarami A 2007. Valuing forests and rangelands-ecosystems services. International Journal of Environmental Research 1: 368-377.
Kawas R, Andrade-Montemayor H and Lu C D 2010 Strategic nutrient supplementation of free-ranging goats. Small Ruminant Research 89: 234-243.
Kaya E and Kamalak A 2012 Potential nutritive value condensed tannin contents of acorns from different oak species. Kafkas Universitesi Veteriner Fakultesi Dergisi, 18(6): 1061-1066.
Kebaili A and Papanastasis V P 2005 Comparison of seasonally available biomass in several woody species of mountainous Mediterranean rangelands. In: A Georgoudis, A Rosati and C Mosconi (Eds) Animal production and natural resources utilisation in the Mediterranean mountain areas. Academic Publishers-bookshop: Wageningen, The Netherlands, pp. 124-128.
Krueger W C 1972. Evaluating animal forage preference. Journal of Range Management 25: 471-475.
Lamy E, Van Harten S, Sales-Baptista E, Mendes-Guerra, M M and Martinho de Almeida A 2012 Factors influencing livestock productivity. In: V Sejian, SMK Naqvi, T Ezeji , J Lakritz and R Lal (Eds) Environmental Stress and Amelioration in Livestock Production. Springer-Verlag: Heidelberg, Berlin, pp.19-33.
Lovreglio R, Meddour-Sahar O and Leone V 2014 Goat grazing as a wildfire prevention tool: a basic review. iForest 7: 260-268.
Lu C D 1988 Implication of forage particle length on milk production in dairy goats. Journal of Dairy Science 70: 1411-1416.
Lynch J J, Hinch G N and Adams, D B 1992 The behaviour of sheep, biological principles and implications for production. CAB International:Wallingford, UK.
Marai I F M, El-Darawany A A, Fadiel A. and Abdel-Hafez M A M 2007 Physiological traits as affected by heat stress in sheep: A review. Small Ruminant Research 71: 1-12.
Mcgregor B A 1992 Nutritional value of some exotic and indigenous plants browsed by goats in southern Australia. Australian Society of Animal Production 19: 307-310.
Meuret M, Bartiaux-Thill N and Bourbouze A 1985 Evaluation de la consommation d’un troupeau de chèvres laitières sur parcours forestier: méthode d’observation directe des coups de dent; méthode du marqueur oxyde de chrome Annales De Zootechnie 34: 159–180.
Min B R and Hart S P 2003 Tannins for suppression of internal parasites. Journal of animal science 81(2): 102-109.
Molina-Alcaide E and Yáñez-Ruiz D R 2008 Potential use of olive by-products in ruminant feeding: A review. Animal Feed Science and Technology. 147: 247-264.
Moore K J and Jung H J G 2001 Lignin and fiber digestion. Journal of range management 54: 420-430.
Narjisse H, Elhonsali M A and Olsen J D 1995 Effects of oak (Quercus ilex) tannins on digestion and nitrogen balance in sheep and goats. Small Ruminant Research 18: 201-206.
NRC 2007 Nutrient Requirements of Small Ruminants. Sheep, Goats, Cervids and New World Camelids. National Academy Press: Washington, DC.
Oosting H J 1956 The Study of plant communities. W. H. Freeman and Co.: San Francisco, California.
Osoro K, Ferreira L M M, García U, Jáuregui B M, Martínez A, García R and Celaya R 2013 Diet selection and performance of sheep and goats grazing on different heathland vegetation types. Small Ruminant Research 109: 119-127.
Pacheco F A L and Van der Weijden C H 2002 Mineral weathering rate calculated from spring water data: a case study in an area with intensive agriculture, the Morais Massif, northeast Portugal. Applied Geochemistry 7: 583-603.
Pacheco L P 2000 A caprinicultura na aldeia de Rouças. Problemática e perspectivas de desenvolvimento. In: G Larcher and H Santos (Eds) Cadernos da Montanha. DRAEDM: Lisboa, pp.132-145.
Papachristou T G and Papanastasis V P 1994 Forage value of Mediterranean deciduous woody fodder species and its implication to management of silvo-pastoral systems for goats. Agroforestry Systems 27: 269-282.
Papachristou T G and Nastis A S 1996 Influence of deciduous broadleaved woody species in goat nutrition during the dry season in Northern Greece. Small Ruminants Research 20: 15-22.
Papanastasis V P 2009 Grazing value of Mediterranean forests. In: M Palahí, Y Birot, F Bravo and E Gorriz (Eds) Modelling, Valuing and Managing Mediterranean Forest Ecosystems for Non-Timber Goods and Services. EFI Proceedings: Joensuu, Finland, pp.7-17.
Pereira E, Castro M and Castro J F 2008 The role of scattered narrow-leafed ash of lameiro systems on livestock production in Northeast Portugal. COST E42, Growing Valuable Broadleaved Tree Species (ValBro).
Pielou E C 1975 Ecological Diversity. Wiley. New York. USA.
Pires J and Moreira N 2000 Meadow pastures: Biomass yield and nitrogen response in the Northeast of Portugal. 11th FAO Meeting of the FAO working group of Mountain Grasslands: Quality and valorisations of mountain animal products. Luz St Sauver, Francia.
Provenza F D, Villalba J J, Dziba L E, Atwood S B and Banner R E 2003 Linking herbivore experience, varied diets, and plant biochemical diversity. Small Ruminant Research 49: 257-274.
Rigueiro A, Silva-Pando F J, Rodríguez R, Castillón P A, Álvarez P, Mosquera R, Romero R and González-Hernández M P 1998 Manual de sistemas silvopastorales. Serie Manuales Técnicos. Universidad Santiago Compostela. Santiago de Compostela. Spain.
Robertson S M 2006 Crop stubbles are as important for sheep production as annual pastures in the Victorian Mallee. Australian Journal of Experimental Agriculture 46: 993-1003.
Rodríguez-Estévez V, García Marínez A, Mata Moreno C, Perea Muñoz J M and Gómez Castro A G 2008 Dimensiones y características nutritivas de las bellotas de los Quercus de la dehesa. Archivos de Zootecnia 57: 1-12.
Rodríguez-Estévez V, García A, Peña F and Gómez A G 2009 Foraging of Iberian fattening pigs grazing natural pasture in the dehesa. Livestock Science 120:135-143.
Sanon H O, Kabore-Zoungrana C and Ledin I 2007 Behaviour of goats, sheep and cattle and their selection of browse species on natural pasture in a Sahelian área. Small Ruminants Research 67: 64-74.
Schlecht E, Dickhoefer U, Gumpertsberger E and Buerkert A 2009 Grazing itineraries and forage selection of goats in the Al Jabal al Akhdar mountain range of northern Oman. Journal of Arid Environments 73: 355-363.
Sequeira E, Aguiar C and Meireles C 2010 Ultramafics of Bragança massif: soils, flora and vegetation. In: N Evelpidou et al (Eds) Natural Heritage from East to West, Springer Verlag: Berlin, Germany, pp.143-149.
Torrano L and Valderrábano J 2005 Grazing ability of European black pine understorey vegetation by goats. Small ruminant research 58: 253-263.
Utsumi S A, Cibils A F, Estell R E, Soto-Navarro S A and Leeuwen D 2009 Seasonal changes in one seed juniper intake by sheep and goats in relation to dietary protein and plant secondary metabolites. Small Ruminant Research 81: 152-162.
Received 10 November 2015; Accepted 7 April 2016; Published 1 May 2016